
Blomst
Blomsten hos en plante er − i den bredeste definition − et uforgrenet skud med begrænset vækst, hvis blade indirekte eller direkte har betydning for den kønnede formering: indirekte som beskyttelses- eller lokkeorganer (blomsterhylster), direkte ved dannelse af forplantningsorganer (støvdragere og frugtanlæg).

1. blomsterbund (receptaculum)
2. bægerblade (sepaler)
3. kronblade (petaler)
4. støvdragere (stamina)
5. frugtanlæg (gynøcium)
En mere snæver definition begrænser begrebet til blomsterplanterne: En blomst hos de dækfrøede består af følgende dele (som dog ikke alle behøver at være til stede): bloster, som kan være differentieret i bæger og krone, støvdragere, frugtanlæg, alle siddende på blomsterbunden. Støvdragerne danner pollen, som ved bestøvningen når frem til frugtanlæggets støvfang. Dér spirer det og danner et pollenrør, som fører sædcellerne frem til ægcellen i frøanlægget. Derved gennemføres befrugtningen, og der udvikles en kim. Frugtanlægget udvikles derefter til én eller flere frugter, som indeholder frø.
Definition
[redigér | rediger kildetekst]
Den brede definition, som ofte ses i den botaniske faglitteratur, omfatter enten udtrykkeligt[1] eller underforstået[2] de nøgenfrøede planter. Det er sjældent, at man stiltiende begrænser definitionen til de dækfrøede under opregningen af blomstens dele.[3] I den tyske standardværk Lehrbuch der Botanik af Eduard Strasburger betragter man blomster som noget, alle frøplanter har, omend med visse begrænsninger: „Hos frøplanterne sidder sporofyllerne næsten altid på kortskud med begrænset vækst. Den slags […] strukturer kan betragtes som blomster, selv om dette begreb oftest bliver brugt om de dækfrøedes tvekønnede sporofylstande og muligvis om nært beslægtede, fossile frøplanter.“[4] Tilsvarende definerer Lexikon der Biologie en blomst helt kortfattet som „et kortskud med begrænset vækst, som bærer sporofyllerne“.[5]
Ordnerne Bennettitales og Caytoniales er fossile repræsentanter uden for frøplanterne, som dannede blomster. Definitionen af ”blomst” svarer ganske vist også til sporofylstandene hos ulvefodsplanter og padderokker[6], men de behandles ikke yderligere her.

I engelsksprogede tekster defineres ”blomst” for det meste mere snævert end det, der svarer til blomster hos de dækfrøede, f.eks. sådan: „et forkortet, kompakt og tvekønnet, reproduktivt skud, som består af megasporangier (frugtblade) og mikrosporangier (støvdragere) samt en steril, egentlig blomst (periant), dannet af mindst ét fladt, sterilt organ.“[7]
Morfologi
[redigér | rediger kildetekst]Hos de dækfrøede er de enkelte organtyper i blomsten ordnet i en fastlagt rækkefølge, regnet nedefra og op, eller, da blomstens akse er forkortet, udefra og ind. Det er også i samme rækkefølge, de bliver skabt ontogenetisk (i løbet af blomsterdannelsen).
Den ”egentlige” blomst
[redigér | rediger kildetekst]Det, som man opfatter som den ”egentlige” blomst (periant), er sterile blade, der dog entydigt tilhører blomsten. Sådan et organ dannes kun hos Gnetopsida og dækfrøede planter. Når alle blomstens blade er ensartede, kaldes de for blosterblade, som f.eks. hos tulipaner. Her kan blostret bestå af en enkelt kreds (monoklamydeisk), to eller flere kredse eller adskillige, skruestillede forløb (homoioklamydeisk). Et dobbelt bloster (heteroklamydeiske blomster) består af uensartede blomsterblade. De ydre, som oftest er grønne, er bægerbladene (sepalerne) og danner svælget (calyx), mens de indre, ofte påfaldende farvede blade, er kronbladene (petalerne), og tilsammen danner de kronen (corolla). Den evolutionsmæssige oprindelse til blomstens blade bliver forstået enten som højblade eller som støvdragere – alt efter slægtskabsforholdene[8].
Mikrosporofyller
[redigér | rediger kildetekst]

Mikrosporofyller er sporangiebærende blade. I sporangierne (pollensækkene) dannes de hanlige mikrosporer. Mikrosporofyller er meget forskelligt udformet alt efter den gruppe af frøplanter, som planten tilhører, og det er ikke helt afklaret, om der består en homologi til de egentlige blade (φύλλον, phýllon = ”blad”). For at undgå den diskussion kunne de også kaldes mikrosporangiebærere. Hos Ginkgo findes mikrosporofyllerne på blomsteraksen, hvor de er skruestillet mange sammen, og hvor de bærer to pollensække ved spidsen. Hos nåletræerne er den hanlige blomst tapformet og består af mange, ofte skruestillede mikrosporofyller, som hver har 2-20 pollensække. Hos Cykas-familien er mikrosporofyllerne skælformede og skruestillede, og de har 5-1.000 pollensække. Hos Welwitschia har de seks sammenvoksede mikrosporofyller hver tre pollensække. Hos Gnetum har det enlige mikrosporofyl en eller to endestillede pollensække. Hos Ledris (Ephedra) er mikrosporofyllet ofte tvedelt og har 2-8 grupper af pollensække, der ofte er sammenvokset to-og-to.
De dækfrøedes mikrosporofyller kaldes støvdragere (stamina). En støvdrager er oftest delt i en støvtråd (filament) og en støvknap (anthere). Den sidste består af to pollensække. Antallet af støvdragere pr. blomst kan variere mellem 1 og 2.000, men er oftest 5-20.[9][10]
Megasporofyller
[redigér | rediger kildetekst]
Megasporofyllerne rummer de frøanlæg, som er typiske for frøplanterne, dvs. strukturer, hvor de hunlige forplantningsorganer er dannet. Også her kunne man bruges den mere neutrale betegnelse megasprangiebærere. Kun hos de dækfrøede betegner man megasporofyllerne som frugtblade.
Hos Cykasplanterne findes frøanlæggene entydigt på blade, og man siger, at de er fyllospore. Det skæl- eller skjoldformede megasporofyl er tydeligt stilket og bærer to frøanlæg ved underkanten af bladribberne (i slægten Cykas dog op til 8). Hos nåletræerne findes der 1-20 frøanlæg på overfladen af et fladt frøskæl, som er et omdannet kortskud. Kun sjældent finder man endestillede frøanlæg på skudspidserne hos nåletræer, men Almindelig Taks er dog et eksempel på det. Tempeltræ danner to frøanlæg for enden af en tvedelt stilk, men hos Gnetopsida dannes der kun ét endestillet frøanlæg pr. blomst.
Hos de dækfrøede er frøanlæggene indesluttet i frugtbladet (karpel), og deraf kommer navnet ”dækfrøede”. Helheden af frugtblade danner tilsammen frugtanlægget (gynoeceum). Antallet af frugtblade pr. blomst ligger på 1-2.000. Oftest er frugtbladet inddelt i en stilkzone og en rørzone, det hule område, hvor frøanlæggene ligger (ovarium)[9]. Frøanlæggene sidder på placenter, og ovenover man ofte finder en griffel, der bærer støvfanget, som er modtagestedet for pollenkornene. Støvfanget er forbundet med frøanlæggene gennem en overførelseskanal eller et tilsvarende væv[11].
Frugtbladene kan være frie (apokarpt eller chorikarpt gynoeceum) eller sammenvoksede (coenokarpt gynoeceum). Hos planter med et sammenvokset frugtanlæg taler man om et stempel (pistil). Sterile blomster har højest nogle reducerede støv- og frugtblade, og de har hyppigt funktion som lokkemidler.
Blomsterakse
[redigér | rediger kildetekst]
På blomsteraksen (blomsterbunden (receptaculum) sidder blomsterbladene. De kan være gestaucht, strakte og mangedobbelte. De kan sidde enkeltvis eller i flere kranse, som er hævet af indskudte vækstpunkter: udformet som gynoeceum (gynofor), gynoeceum og androeceum (androgynofor), eller gynoeceum, androeceum og krone (anthofor).
Der kan også være ringformede områder, som er hævet i vejret omkring centrum. Et eksempel er det stamen-corollarør, som er typiske hos unge kurveblomster, eller det bægerformede svælg hos mange planter i Græskar-familien. En udbredt blomsterbund kaldes et blomsterbæger (hypanthium).
Når alle dele ud over frugtknuden er hævet i vejret, opstår der et blomsterrør, som bærer blomsterhylsteret og støvdragerne. Det kaldes et omkringsædigt blomsterbæger. En oversædig frugtknude sidder frit på blomsteraksen. Når ydersiden af frugtbladene er trukket med ind i blosterbægeret, dannes der en undersædig frugtknude.
Nektarier
[redigér | rediger kildetekst]Nektarier er kirtler, der udskiller nektar som lokkemad for de bestøvende insekter. Den består mest af en sukkeropløsning, som dog også kan indeholde aminosyrer og vitaminer. Det er mere sjældent, at der dannes olier som f.eks. hos Fredløs (Lysimachia). Nektarier kan være dannet af blomsterhylstret, støvdragerne, frugtknuden eller blomsteraksen, men også uden for den egentlige blomst.
Blomsterstilling og placering
[redigér | rediger kildetekst]Blomsterbladene kan – alt efter bladstillingen – være anbragt skruestillet (acyklisk) eller kransstillet (cyklisk). En skruestillet placering bliver ofte betragtet som et primitivt træk, og det er ofte forbundet med et stort antal blomsterblade. Vinklen mellem to blade, der følger efter hinanden, er ofte tæt på det gyldne snit på ca. 137,5°.
Det er ofte, at kun en del af blomsterbladene er skruestillede, mens de indre blomsterblade er ordnet i kranse. Den slags blomster bliver betegnet som spirocykliske eller hemicykliske. En overgangsform mellem skruestillet og kransstillet opstår, når der indskydes en pause i bladstillingen efter hver tre blade som hos Yulan-Magnolia (Magnolia denudata), sådan at der opstår en 1-3tallig krans. Også hos mange kransstillede blomster kan man skimte en skrueformet bladstilling under udviklingen, men bladene i en kreds står dog så tæt sammen, at der opstår en krans.
Hos de kransstillede findes der kun én type blomsterblade i hver kreds eller krans. Det passer også med denne stilling, at der opstår en reduktion i antallet af blomsterblade (oligomere kredse). De fleste kransstillede, tvekønnede blomster har en fire- (tetracyklisk) eller femtallig (pentacyklisk) kransopbygning: En bægerblads-, en kronblads-, en eller to støvdrager- og en frugtbladskrans. Hos de fleste dækfrøede er antallet af blade pr. krans fastlagt, og man taler så om to-, tre-, fire- eller femtallige blomster. Hvis alle kredse består af det samme antal, kaldes blomsten isomer (som hos Tulipan), men hvis det ikke er tilfældet, kaldes blomsten heteromer.
Bladene i de kranse, der er oven over hinanden, er for det meste anbragt ud for mellemrum (alternansprincip), dvs. at medlemmerne af de næstinderste kranse sidder i mellemrummet mellem to medlemmer af den foregående krans. Hos de pentacykliske blomster opstår ofte den situation, at den indre støvdragerskreds bliver trængt udad af frugtknuden og tilsyneladende bliver til den yderste. Dette fænomen kaldes obdiplostemoni.
Alle blomsterblade kan være mere eller mindre sammenvoksede enten indbyrdes eller med andre.
Når der kan trækkes flere end tre symmetriplaner gennem en blomst, så er den radiærsymmetrisk (som hos Tulipan). Med to symmetriplaner er den disymmetrisk, som f.eks. hos Dicentra. Et enkelt symmetriplan findes hos zygomorfe (dorsiventral) blomster. Oftest ligger dette symmetriplan i blomstens median (og forløber altså gennem støttebladet og dets skudakse), som hos de læbeblomstrede, mere sjældent vinkelret på medianeplanet som hos Lærkespore. Asymmetriske blomster har ingen symmetriplaner. Strengt talt kan man også regne skruestillede blomster med til gruppen, selv om de ofte bliver anset for at være radiærsymmetriske.
Blomsterdelenes forhold til hinanden kan vises grafisk i blomsterdiagrammer eller i blomsterformler.
Ofte er flere eller mange enkeltblomster forenet i en blomsterstand.
Kønnethed
[redigér | rediger kildetekst]Hos de dækfrøede er den tvekønnede blomst den oprindelige form. ved siden af findes der enkønnede blomster, hvor organerne fra det undertrykte køn dog oftest findes som rudimenter: Der findes blomster, som kun viser støvdragere (”hanlige”), og andre, som kun har støvfang (”hunlige”). Planter og slægter med énkønnede blomster kan være sambo, hvor hanlige og hunlige blomster findes på samme individ, eller særbo, hvor de hanlige og hunlige blomster findes på forskellige individer. Endelig er der de såkaldt flerbo planter, hvor der findes både tvekønnede, hanlige og hunlige blomster på ét og samme individ.
Blomsterinduktion
[redigér | rediger kildetekst]Blomsterinduktion er omdirigeringen af plantens vækst fra vegetativ skud- og bladvækst til generativ vækst med blomsterdannelse. De udløsende faktorer kan være indre (endogene) eller ydre (exogene). Induktionen er uigenkaldelig og kan altså ikke vendes eller standses. Der findes fire vigtige udløsende elementer:
- Endogene eller autonome udløsere kan være til stede, selv om man ikke kender noget nærmere til deres natur. Hos mange arter er en bestemt alder eller en bestemt størrelse en nødvendig forudsætning for, at de kan blomstre, sådan som det kendes fra mange træarter. Hos have-Ært er blomstringstidspunktet fastlagt i arveanlæggene hos den enkelte sort. Sentblomstrende økotyper af Almindelig Gåsemad (Arabidopsis thaliana) sætter blomst fra og med en bestemt alder og af egen drift, hvis der ikke forud har været nogen induktion fra ydre miljøfaktorer. Omdirigering til blomsterdannelse sker efter en aktivering af bestemte gener som f.eks. FCA (Flowering Control Arabidopsis).
- Hos de fleste planter bliver blomstringstidspunktet fastlagt ved ydre faktorer. De vigtigste faktorer er i den forbindelse daglængden og kuldepåvirkning (vernalisation). Daglængden har betydning for både kortdagsplanter og langdagsplanter, der er afhængige af et bestemt antal lystimer pr. døgn. I den forbindelse spiller bladenes fytokromsystem en vigtig rolle sammen med cryptokrom. Proteinet FKF1 tjener som fotonmodtager og måler daglængden. Dette system påvirker det såkaldte ”konstansgen”, hvis aktivering i sig selv er tilstrækkelig til at udløse blomsterinduktionen. Man har længe forventet at finde et blomsterhormon, ”florigen”, der kan tjene som signal fra bladene, men det forblev længe ukendt. Genet FT (se nedenfor) betragtes nu som et muligt emne, da det bliver aktiveret i bladene, men først virker i blomstervækstpunkterne.[12][13]
- Særligt de monokarpe planter (som kun blomstrer én gang) har brug for en kuldepåvirkning, dvs. en længere kuldeperiode, for at kunne komme i gang med blomstringen. Blandt de gener, som bliver aktiveret ved kulde, findes Frigida (FRI) hos Arabidopsis samt VRN1 og VRN2 hos vinterhvede. Deres genprodukter hæmmer genet FLC, som på sin side undertrykker blomsterdannelsen. FRI eller VRN muliggør blomsterdannelsen, men udløser den ikke. Dertil skal ofte bruges flere andre, vigtige faktorer.
- Bestemte plantehormoner, som f.eks. gibberellinerne, er vigtige for stimulering af blomstring, når langdagsforhold ikke findes, men den egentlige virkningsmekanisme er dog ukendt[14].
Blomsterdannelse
[redigér | rediger kildetekst]Når et meristem er omdannet fra et vegetativt til et generativt vækstpunkt, sættes den egentlige blomsterdannelse i gang af en lang række, indbyrdes forbundne regulatorgener.
Symmetrigener
[redigér | rediger kildetekst]De såkaldte symmetri- eller Katastergener fastlægger blomstens struktur, dens symmetriplaner, antallet af blomster og deres organer. Mutationer i disse gener ændrer blomstens arkitektur, og generne bliver ofte benævnt efter de fænotyper, der bliver fremkaldt af mutationerne: Mutationer i generne cycloidea (CYC) eller dichotoma (DICH) hos Løvemund skaber radiærsymmetriske blomster i stedet for de sædvanlige, zygomorfe. Disse mutationer optræder også naturligt og kaldes pelori. En mutation i genet cauliflower (CAL) hos Gåsemad og hos Broccoli skaber mange forgreninger i blomsterstanden uden brugbare blomster. CAL hæmmer genet TFL1, som er et skudidentitetsgen, der fremmer forgreningen i blomsterstanden. tfl1-mutanter danner på tilsvarende måde få- eller énblomstrede blomsterstande hos Arabidopsis.
Blomstringsgener
[redigér | rediger kildetekst]
Inden i den blomsterstand, der er ved at opstå, bliver meristemer til blomstermeristemer, når generne Leafy (LFY) og Apetala1 (AP1) bliver aktiveret. Begge koder for transkriptionsfaktorer og tilhører en gruppe af gener, der integrerer blomstringssignaler. Viden om disse og de fleste andre gener blev skaffet ved undersøgelser af Gåsemad, Løvemund og en Petunia-hybrid. De vigtigste integrationsgener er:
- FLC (Flowering Locus C) integrerer signaler fra kuldepåvirkning og autonome signaler. FLC er en transkriptionsfaktor, og når den bliver bremset, opstår der tidlig blomstring. Mængden af FLC-protein/mRNA svarer nøje til blomstringstidspunktet. FLC aktiverer blomstringsaktivatoren SOC1.
- SOC1 (Suppressor of Overexpression of Constans) bliver aktiveret af gibberelliner via CO af langdagspåvirkning og FLC. Ophævelse af undertrykkelsen via FLC er ikke tilstrækkeligt til at aktivere SOC1: Der skal yderligere ske en aktivering via CO eller gibberelliner.
- LFY bliver ligesom SOC1 aktiveret via gibberelliner og langdagspåvirkning. Det sidste sker ved hjælp af SOC1 eller ved hjælp af Agamous-Like24 (AGL24).
- FT (Flowering Locus T) bliver aktiveret via langdagspåvirkning og ved hjælp af CO, der ellers er holdt undertrykt af FLC. FT aktiverer desuden AP1.
De to sidste gener i rækkefølgen er LFY og AP1, de såkaldte blomstermeristem-identitetsgener, som dog påvirker hinanden: AP1 bliver aktiveret af LFY, men der bliver under alle omstændigheder dannet blomst, også selv om LFY mangler ganske vist uden de rigtige organer. Det førnævnte TFL1 aktiverer LFY og AP1, som på den anden side aktiverer hinanden. TFL1 tjener til at forhindre et alt for tidlig blomstring.
Fastlæggelse af blomstens organer
[redigér | rediger kildetekst]
Blomsterorganerne bliver fastlagt efter ABC-modellen. Det er tre klasser af gener, som bestemmer blomsterorganernes identitet via gensidige påvirkninger (organidentitetsgener):
- A fører til dannelse af bægerblade
- A og B fører til dannelse af kronblade
- B og C fører til dannelse af støvdragere
- C fører til dannelse af frugtblade
A og C undertrykker gensidigt hinanden, sådan at det ikke kommer til Überschneidung. Når den ene mangler, bliver den anden klasse skabt i hele blomsten. ABC-modellen blev opstillet af E. Coen og E. Meyerowitz[15], og den er i de mellemliggende år blevet bekræftet og udvidet ad flere omgang.
Hos Arabidopsis kendes til dato følgende gener:
- Klasse A: AP1 og AP2
- Klasse B: AP3 og Pistillata (PI)
- Klasse C: Agamous (AG)
Disse gener bliver udtrykt vedvarende i de organer, som de styrer, og ikke blot i begyndelsen. AP1 er altså ikke kun et blomstermeristemgen, men også et organidentitetsgen. I blomsterdannelsens tidlige fase bliver det dannet i hele meristemet, men senere kun i blomstens organkredse. Mutationer fører til skift i organernes identitet. Når A mangler, udvikles der frugtblade i stedet for bægerblade og støvdragere i stedet for kronblade (fordi C bliver dannet). Tredobbelte mutanter danner kun normale blade. C-mutanter har ingen begrænsning i væksten, og meristemet vokser videre uden grænser.
Senest har man opdaget en fjerde klasse, E, som består af de fire gener Sepallata1 til 4 (SEP) hos Gåsemad. De bliver aktiveret i kron-, støv- og frugtbladene, og de tager sig også af organidentitet. Triple-mutanter (sep1 til 3) danner kun bægerblade, Firedobbelte mutanter kan kun danne løvblade.[16]
En femte klasse, D, består af gener, der er ansvarlige for frøanlæggenes identitet. Hos Petunia er det generne Floral Binding Protein (FBP) 7 og 11. Doppeltmutanter danner frugtbladagtige strukturer i stedet for frøanlæg. Hos Gåsemad hedder de tilsvarende gener Seedstick (STK), Shatterproof1 (SHP1 og SHP2)
Den overvejende del af blomsterorgangenerne tilhører familien af MADS-Box-gener. ABCE-generne er nødvendige både til dannelsen af blomstens organer ved udformningen af vegetative organer. Dette har man kunnet vise ved aktivering af generne uden for deres normale placering.
Det er først i disse år ved at blive afklaret, hvordan blomteridentitetsgenerne, der bliver aktiveret i hele blomstermeristemet, kan styre organidentitetsgenerne. Det ser ud til, at B-Klasse-genet AP3 som det første bliver aktiveret ved kombinationen af LFY, AP1 og genet Unusual Flower Organs (UFO). LFY aktiverer AG, C-Klasse-genet, sammen med en yderligere koaktivator, Wuschel (WUS). Derefter undertrykker AG på sin side WUS, og derved bliver væksten i blomsteraksen hæmmet.
Theissens kvartetmodel fra 2001 har været endnu et skridt mod en forståelse af disse mekanismer. Den hævder, at blomsterorganiserings-proteinerne virker i tetramerer, dvs. som kvartetter. For hvert organ findes skal der med andre ord fiindes mindst ét specifikt tetramer. Man har foreløbig kunnet vise, at hos gær danner proteinerne virkelig tetramerer. Modellen tjener allerede som arbejdsgrundlag for mange undersøgelser.[16]
ABC-modellens mange varianter
[redigér | rediger kildetekst]
Sammenlignende undersøgelser hos en række plantearter har vist, at ABC-modellen i den beskrevne form kun gælder for gruppen Eudikotylae. Især hos den basale gruppe af dækfrøede findes der forskellige udformninger. Hos Åkande (Nuphar) er der ikke nogen skarp grænse for aktivering af generne (de har “fading borders”), og det betyder, at der findes en glidende overgang mellem bægerbladsagtige blosterblade over kronbladsagtige blosterblade til Staminodien, støvdragere og frugtblade.[16]
Tulipan har et ensartet blomsterhylster i to kredse. Her bliver B-klasse-generne aktiveret i begge kredse.[16]
Blomsterne hos græserne er meget stærkt ændrede. Alligevel viser ABC-modellen, hvordan foravner (Palea) bliver reguleret af A-generne, mens svulmelegemer (Lodiculae) reguleres af A og B, altså svarende til forholdene hos bæger og krone.[16]
Skræppe (Rumex) danner et ensartet, men reduceret og uanseligt blomsterhylser. Her er B-klassen begrænset til støvdragerne, og derfor bliver både bæger og krone reguleret af A-generne.[16]
-
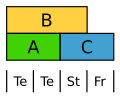 ABC-model fo Tulipan. Te: blosterblade
ABC-model fo Tulipan. Te: blosterblade -
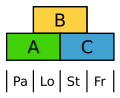 ABC-model hos Ris. Pa: foravne, Lo: svulmelegeme
ABC-model hos Ris. Pa: foravne, Lo: svulmelegeme -
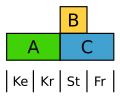 ABC-model hos Skræppe
ABC-model hos Skræppe



Imidlertid betvivler man A-klassens almengyldighed som identitetsgener for blomsterorganerne, og A-klassegener spiller som nævnt også en rolle ved fastlæggelsen af blomstermeristemets identitet. Det har kun kunnet påvises hos Gåsemad, at de har betydning for dannelsen af blomsterorganerne, og de spiller måske kun en rolle hos de nærtbeslægtede. Hos Løvemund er BC-generne tilstrækkelige til blomsterdannelsen, men det står hen, om det gælder for andre grupper blandt de dækfrøede.[17]
Blomstring (antese)
[redigér | rediger kildetekst]Blomstringsforløbet bliver betegnet som antese.
Blomsteråbning
[redigér | rediger kildetekst]Forud for åbningen af blomsten, danner den en knop, der svarer til overvintringsknopperne. Her tjener bægerbladene som beskyttelse. Åbningsproessen kan gennemføres via forskellige mekanismer:
- Reversibel ophobning af ioner. Det er påvist entydigt hos Stængelløs Ensian (Gentiana kochiana).
- Programmeret celledød i afgrænsede områder af blomsterbladene.
- Vandtab om dagen og genopfyldning i løbet af natten. Det findes bl.a. hos Aften-Pragtstjerne (Silene latifolia), som er natblomstrende, og hvis kronblade rulles ind om dagen som følge af væsketab.
- Forskellig væksthastighed på yder- og indersiden af blomsterbladene. Hos Tulipan har indersiden af blomsterbladene et ca. 10 °C lavere vækstoptimum end ydersiden, og det fører til en åbning om morgenen og en lukning om aftenen.
En vigtig faktor for udspilingen af kronblade er forøgelsen af turgortrykket, som mest opstår ved en øget sukkerkoncentration efter nedbrydning af højmolekylære kulhydrater (f.eks. stivelse hos Roser eller fruktaner hos Dagliljer). Deraf følger en udvidelse af cellevæggen, men opgivelserne over plantehormonernes rolle er modstridende i litteraturen.[18] De cellefysiologiske og særligt de genetiske baggrunde for blomsternes åbning og lukning er knap nok kendt endnu. Blomstens åbning bliver udløst af ydre faktorer. Hos natblomstrende arter er det ofte den forøgede luftfugtighed om aftenen, der er afgørende. Derimod reagerer især de forårsbomstrende på højere temperaturer (Almindelig Vintergæk (Galanthus nivalis), Krokus-arter (Crocus spp.)). En tredje gruppe reagerer på lys, som f.eks. Almindelig Tusindfryd (Bellis perennis). De længelevende blomsters åbning og lukning foregår ved de samme mekanismer, men er også underkastet en indre rytmik.
Lukningen af blomsten kan foregå ved vækstforskydninger eller ved reversible turgorændringer. I disse tilfælde er der grundlag for en gentagen åbning og lukning. Tab af turgortryk på grund af aldring fører til en vedvarende lukning af blomsten.[19]
Bestøvning
[redigér | rediger kildetekst]
Bestøvningen er overførslen af den hanlige pollen til de hunlige modtageorganer: Mikropylen hos de nøgenfrøede og støvfanget hos de dækfrøede. Bestøvningen er derfor ikke identisk med befrugtningen. Bestøvning kan foregå ved hjælp af pollen fra samme individ (Selvbestøvning, Autogami), eller med pollen fra et andet individ (Fremmedbestøvning, Allogami).
Selvbestøvning nedsætter dog den genetiske variation, og derfor findes der forskellige tilpasninger hos planterne, som skal hindre selvbestøvning eller selvbefrugtning:
- Herkogami er den rumlige adskillelse af støvknapper og støvfang, sådan at selvbestøvning ikke er mulig. Hos Kapers (Capparis spinosa) er hele frugtknuden hævet på en måde, så støvfanget står over støvknapperne i insekternes indflyvningsbane.
- Dichogami er den tidsmæssige adskillelse af støvknappernes og frugtknudens modning. Derfor findes der førsthanlige (proterandri) og førsthunlige blomster (proterogyni). Samtidig modning kalder man homogami.
Dichogami og herkogami kan ganske vist hindre bestøvning inden for den samme blomst, men ikke mellem forskellige blomster på samme plante (geitonogami). Derfor har mange arter udviklet yderligere undvigelsesmekanismer:

- Selvsterilitet: Her bliver selvbefrugtning forhindret ved genetiske faktorer. Hyppigt kan man også se den slags spærringssystemer i plantens udformning (heteromorfi): et velkandt eksempel er heterostyli hos Kodriver (Primula).
- Bestøvingsmetoder
Alt efter bestøverens type findes der tre store tilpasningsmåder: vindbestøvning (anemofili), vandbestøvning (hydrofili) og dyrebestøvning (zoofili). De nøgenfrøede er hovedsagelig vindbestøvede, mens de første dækfrøede sandsynligvis var dyrebestøvede. Først i anden omgang har der udviklet sig vind- eller vandbestøvning hos flere slægter blandt de dækfrøede. De vigtigste kendetegn ved de enkelte bestøvningstyper er:
- Anemofili (vindbestøvning): uanselig blomst; reduceret antal blomsterorganer (fx bægerblade og kronblade); særbo eller sambo blomster findes ofte; tætte og ofte hængende blomsterstande; meget lidt eller ingen pollenkitt; glat pollenoverflade; støvfang med stor overflade; ét eller få frøanlæg pr. blomst; ingen nektar.
- Hydrofili (vandbestøvning): uanselig blomst; særbo eller sambo blomster findes ofte; opbygning af luftvæv; vandtætte pollenvægge; trådformede pollenkorn; støvfang med stor overflade; ét eller få frøanlæg pr. blomst.
- Zoogami (dyrebestøvning): Hos de dyrebestøvede planter er det ikke blomsten som morfologisk enhed, men blomsten som funktionel enhed, der står i forgrunden. Her svarer ”blomsten” ofte til en enkelt blomst (Tulipan), men hyppigt er mange, enkelte blomster samlet til en ”blomst”-lignende dannelse, som kaldes et pseudantium. Eksempler findes hos de kurveblomstrede og de skærmblomstrede. Det sker mere sjældent, at en enkeltblomst danner flere ”blomster”, sådan som det sker hos Iris (Meranthium). De vigtigste kendetegn på dyrebestøvede blomster er: Tvekønnede blomster eller ”blomst”agtige stande; dækfrøethed; påfaldende farve og/eller duft; pollen og/eller nektar som næringstibud (eller snydeindretninger); stærkt skulpterede pollenoverflader og meget pollenkitt.
Befrugtning
[redigér | rediger kildetekst]Hos de nøgenfrøede når pollenkornene frem til frøanlæggets mikropyle. For det meste trækkes de ind i pollenkammeret efter en indtørring af bestøvningsdråben. Afhængigt af planteslægten vil der så enten frigives en spermatozoid i pollenkammeret eller også vil pollenrøret spire frem. Der kan gå op til seks måneder mellem bestøvning og befrugtning (visse arter af Cykas-familien).
Hos de dækfrøede når pollenkornet frem til støvfanget. Pollenrøret når frem til frøanlæggene gennem et specielt ledevæv. Hvor der findes et ensartet væv i griflen, sådan at pollenrørene fra et frugtblad kan føre frem til et andet frugtblad, kalder man ledevævets helhed for et compitum.
Når pollenrørene henholdsvis spermatozoiderne er nået frem til ægcellerne, sker den egentlige befrugtning. Hos de dækfrøede, hos Gnetum og hos Ephedra foregår der en dobbelt befrugtning: Hos de dækfrøede smelter én af de to spermakerner sammen med ægcellen og danner en Zygote. Den anden smelter sammen med en af de diploide kimsækkerner til en triploid endospermkerne, som bliver til frøets næringsvæv (endosperm). Hos Ephedra smelter den anden spermakerne sammen med arkegoniets kanalcelle, hos Gnetum smelter de to spermaceller med to gametofytceller, men der udvikles for det meste kun én af de to Zygoter.
Efter befrugtningen udvikles zygoten til en kim (embryo), frøanlæggene til frø, og blomsten til en frugt.
Ældning
[redigér | rediger kildetekst]Blomsterne er planternes forplantningsorganer, og der dannes hele tiden nye, modsat hvad der er tilfældet hos dyrene. Livslængden er nøje afmålt, for blomsterne bruger mange ressourcer, og frugtanlægget er en væsentlig adgangsvej for sygdomskim. Blomster, som allerede er bestøvet, skal heller ikke konkurrere unødigt med de endnu ikke besøgte om bestøvernes interesse.
Bestøvningen med pollen er en vigtig udløser for blomstens ældning, og hos de fleste arter vil det forkorte blomstens livslængde betragteligt. Hos visse planter udløser bestøvning dannelsen af plantehormonet ethylen, og det fremkalder på sin side ældning hos kronbladene. Andre arter er ikke følsomme over for etylen, og deres ældningsmekanisme er ukendt. Efter bestøvningen dør krone, støvdragere og griffel, mens frugtknuden udvikler sig videre til en frugt. Denne bortdøen går hånd i hånd med en tilbagetrækning af vigtige stoffer fra disse organer, ganske som det sker hos blade før løvfald.[20]
Evolution
[redigér | rediger kildetekst]Den tidlige udvikling af de dækfrøede planters blomst er ikke afklaret, da der næsten ikke findes fossile forløbere for de ”moderne” versioner. Ligeledes er slægtskabsforholdet til de andre frøplanter endnu uklart, og der findes to forskellige hypoteser til forklaring af blomsternes opståen. Man antager, at dannelsen af tvekønnede blomster hos de dækfrøede kan være opstået ved tilpasning til biller som bestøvere. Omplaceringen af frøanlæggene til lukkede frugtblade kunne så være en beskyttelse mod billernes gnav.
Arber og Parkins euantiteori fra 1907[21] går ud fra, at de dækfrøedes forgængere allerede havde tvekønnede blomster, og at disse blomster er et enakset system med sidestillede mikro- og megasporofyller. Støv- og frugtbladene er med andre ord homologe med løvblade. Denne teori hviler på analyser af den fossile slægt Caytonia, der anses for at være en mulig forløber for de dækfrøede planter. Caytonia havde fjerdelte mikro- og megasporofyller, som muligvis var samlet i en tvekønnet blomst. I megasporofyllet sad cupulae med indtil flere frøanlæg sidestillet på en rhachis. Mikrosporofyllerne var ligeledes fjerdelte, og hver fjer bar flere grupper af sammenvoksede pollensække. Frugtbladet hos de dækfrøede kan være opstået som en udfladning af rhachis, mens støvbladet er dannet som en reduktion til en enkelt stilk med to sammenhængende støvknapper á to pollensække.
Richard Wettsteins pseudantiteori[22] antager, at blomsten hos de dækfrøede er opstået ud fra en bomsterstand af énkønnede blomster. Derfor må støvblade og frugtanlæg oprindeligt have været sideskud og ikke blade. Grundlaget for denne teori er den antagelse, at de dækfrøede nedstammer fra Gnetopsida. Frugtanlægget opstod dermed ud fra en omdannelse af blomstens dækblad, mens andre dele af frøanlægget blev dannet på grundlag af højblade under blomsten. Denne teori bliver dog næsten ikke støttet af molekylære og morfologiske analyser.
Selv om de fleste forfattere nu hælder til euantiteorien, er dannelsen af støvbladet, frugtbladet og frøanlæggets andet integument stadigvæk uklart forstået.
Molekylærbiologisk begrundede teorier
[redigér | rediger kildetekst]Blomsterne hos de ældste dækfrøede var tvekønnede og havde et udifferentieret blomsterhylster (”blomsten”) med organerne i spiralstilling eller med mere end to kranse.[16]
Bortset fra frøplanterne findes der ingen blomsterorgan-identitetsgener eller bare noget, der svarer til dem. Hos de nøgenfrøede, der kun danner enkønnede blomster, findes der gener, der svarer til klasse B og C. Deres aktivering ligner den hos de dækfrøede: C er aktiv i alle reproduktive organer, men B kun i de hanlige blomster. Baseret på disse erkendelser har man opstillet flere hypoteser om opkomsten af den tvekønnede blomst hos de dækfrøede:
- Ifølge den “out of male”-hypotese, der blev opstillet af Theissen m.fl. i 2002, dannede de hanlige blomsterkogler hunlige organer efter en reduktion af B-klasseaktiveringen i den øverste del af koglen. Efter “out of female”-hypotesen blev der dannet hanlige organer ved den nederste del af hunlige kogler. Et blomsterhylster opstod først efter tvekønnetheden ifølge begge disse modeller.[16]
- ”Mostly male”-hypotesen, der først blev foreslået i 2000 af Frohlich og Parker, går også ud fra den hanlige blomst.[23] De opdagede, at der forekommer to udgaver af Leafy-genet. Leafy styrer dannelsen af hanlige, mens genet Needly styrer dannelsen af hunlige blomster. Needly findes hos alle nøgenfrøede bortset fra Gnetum, men ikke hos de dækfrøede. Teorien siger nu, at de hanlige kogler blev tvekønnede ved en deaktivering af Needly-genet.[24][16]
Udviklingen af de dækfrøedes blomster skete via fire nøglehændelser:[24]
- Udvikling af den tvekønnede blomsterakse
- Udvikling af den nedsænkede blomsterakse og begrænsning af dens vækst: Dette skete ved hjælp af C-klassegenerne, som undertrykker Wuschel, meristemets vedligeholdelsesgen.
- Udvikling af et blomsterhylster med kronblade
- Udvikling af den klassiske, todelte blomst med bæger og krone.
Soltis m.fl. (2007) diskuterer den hypotese, at ABC-modellen, der kendes fra Gåsemad, ikke fandtes hos de første dækfrøede, men derimod et system med uklare grænser som hos Nøkkerose. Ud fra den grundmodel kan ABC-modellen være opstået ganske simpelt ved forskellige markeringer af grænserne, ligesom også de forandrede former som f.eks. hos Tulipan kan afledes derfra.[17]
Brugen af blomster
[redigér | rediger kildetekst]
I modsætning til de frugter, der kommer af blomsterne spiller blomster en underordnet rolle for menneskers ernæring. Blomstestandene hos Blomkål og Artiskok bliver brugt som gemyse, mens Hampeplantens bliver brugt som rusmiddel. Nogle blomster eller blomstestande bliver brugt som salat eller som maddekoration (f.eks. Marts-Viol, Hjulkrone og Tallerkensmækker). Blomster, knopper, blomsterdele eller hele blomstestande bliver brugt som krydderi: Lavendel (Lavandula angustifolia), Kryddernellike (Syzygium aromaticum), Safran (Crocus sativus) og Almindelig Humle (Humulus lupulus).[25]
Blomsterne spiller dog en væsentligt større rolle som pynt i form af prydplanter og snitblomster. Morgenrødens gudinde (Aurora/Eos) strøer blomster som symbol på den nye dags lysstråler. Blomster er også kendetegnende for gudinden Flora. Blomsterne er knyttet til foråret, og til lugtesansen. Blomster har siden antikken været tegn på skønhedens og livets forgængelighed. I de malede ”nature morte” ses altid en vissen blomst eller nogle affaldne blomsterblade som symbol på Vanitas. I logikken fremstilles allegorien til håbet ofte med blomster i armen for at minde om, at der snart vil udvikles en frugt af blomsten.[26] Enkelte arter har deres egne, symbolske betydninger, som dog kan ændres i århundredernes løb, selv om rosen er og bliver symbol på Venus, Dionysos og Maria, og symbolet på den jordiske og guddommelige kærlighed, for jomfruer såvel som for prostituerede.[27]
Kilder
[redigér | rediger kildetekst]Artiklen er hovedsageligt baseret på følgende kilder:
- Peter Leins: Blüte und Frucht. Morphologie, Entwicklungsgeschichte, Phylogenie, Funktion, Ökologie. E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart 2000, ISBN 3-510-65194-4
- P. Sitte, E. W. Weiler, J. W. Kadereit, A. Bresinsky, C. Körner: Strasburger – Lehrbuch der Botanik für Hochschulen. 35. Auflage, Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X
Afsnittet om blomsterinduktion og blomsterdannelse bygger på:
- Peter Schopfer, Axel Brennicke: Pflanzenphysiologie. Elsevier, München 2006. Afsnittet Blütenbildung und Befruchtung, S. 501–523, ISBN 978-3-8274-1561-5
- Thomas Jack: Molecular and Genetic Mechanisms of Floral Control. The Plant Cell, Band 16, S. S1–S17, Supplement 2004.
- Paul K. Boss, Ruth M. Bastow, Joshua S. Mylne, Caroline Dean: Multiple Pathways in the Decision to Flower: Enabling, Promoting, and Resetting. The Plant Cell, Band 16, S. S18–S31, Supplement 2004.
Noter
[redigér | rediger kildetekst]- ^ Gerhard Wagenitz: Wörterbuch der Botanik. 2. Auflage, Spektrum Akademischer Verlag, Heidelberg, Berlin 2003, S. 47f. ISBN 3-8274-1398-2; Schück, Schuck, Stimm: Lexikon der Baum- und Straucharten. Nikol, Hamburg 2002, S. 83. ISBN 3-933203-53-8
- ^ Manfred A. Fischer (Red.): Exkursionsflora für Österreich, Liechtenstein und Südtirol. Oberösterreichisches Landesmuseum, Linz 2005, S. 90. ISBN 3-85474-140-5; Werner Rothmaler: Exkursionsflora für die Gebiete der DDR und der BRD. Band 2: Gefäßplanten. 13. Auflage, Volk und Wissen VEB, Berlin 1987, S. 24. ISBN 3-06-012557-0
- ^ Siegmund Seybold (Hrsg.): Schmeil-Fitschen interaktiv (CD-Rom), Quelle & Meyer, Wiebelsheim 2001/2002, ISBN 3-494-01327-6
- ^ Strasburger 2002, S. 750.
- ^ Stikordet „blomst“ i: Herder-Lexikon der Biologie. Spektrum Akademischer Verlag GmbH, Heidelberg 2003. ISBN 3-8274-0354-5
- ^ Strasburger 2002, S. 720, 723; Wagenitz: Wörterbuch der Botanik 2003, S. 47.
- ^ “a determinate, compressed, bisexual reproductive axis composed of megasporangia (carpels), microsporangia (stamens) and a sterile perianth composed of at least one sterile laminar organ”. G. Theißen, R. Melzer: Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower. Annals of Botany, Band 100, 2007, S. 603–619.; det svarer ret nøje til en definition, der bygger på et overblik over litteraturen, og som findes i: Richard M. Bateman, Jason Hilton, Paula J. Rudall: Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers. Journal of Experimental Botany, Band 57, 2006, S. 3471–3503.
- ^ Lore Steubing og Hans O . Schwantes: Ökologische Botanik, 1992 ISBN 978-3825208882 side 233-236 (tysk)
- ^ a b Lore Steubing og Hans O . Schwantes: Ökologische Botanik, 1992 ISBN 978-3825208882 side 242 (tysk)
- ^ E.J.H. Corner: The Life of Plants, 2002, ISBN 978-0226116150 side 194 (engelsk)
- ^ E.J.H. Corner: The Life of Plants, 2002, ISBN 978-0226116150 side 195 (engelsk)
- ^ Katja E. Jaeger, Alexander Graf, Philip A. Wigge: The control of flowering in time and space. Journal of Experimental Botany, Band 57, 2006, S. 3415–3418.
- ^ Laurent Corbesier, George Coupland: The quest for florigen: a review of recent progress. Journal of Experimental Botany, Band 57, 2006, S. 3415–3418.
- ^ Lore Steubing og Hans O . Schwantes: Ökologische Botanik, 1992 ISBN 978-3825208882 side 228 (tysk)
- ^ E. S. Coen, E. M. Meyerowitz: The war of the whorls: genetic interactions controlling flower development. Nature Band 353, S. 31–37.
- ^ a b c d e f g h i G. Theißen, R. Melzer: Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower. Annals of Botany, Band 100, 2007, S. 603–619.
- ^ a b D. E. Soltis, A. S. Chanderbali, S. Kim, M. Buzgo, P. S. Soltis: The ABC Model and its Applicability to Basal Angiosperms. Annals of Botany, Band 100, 2007, S. 155–163.
- ^ Se en oversigt hos Wouter G. van Doorn og Uulke van Meeteren: Flower opening and closure: a review, 2003.
- ^ Der Abschnitt beruht auf: Wouter G. van Doorn og Uulke van Meeteren: Flower opening and closure: a review, 2003.
- ^ H. J. Rogers: Programmed Cell Death in Floral Organs: How and Why do Flowers Die? Annals of Botany, Band 97, 2006, S. 309–315.
- ^ E. A. N. Arber, J. Parkin: On the origin of the angiosperms. Botanical Journal of the Linnean Society, Band 38, 1907, S. 29–80.
- ^ Richard Wettstein: Handbuch der Systematischen Botanik, Band 2. Wien, Deuticke 1907.
- ^ M. W. Frohlich, D. S. Parker: The mostly male theory of flower evolutionary origins: from genes to fossils. Systematic Botany, Band 25, 2000, S. 155–170.
- ^ a b Richard M. Bateman, Jason Hilton, Paula J. Rudall: Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers. Journal of Experimental Botany, Band 57, 2006, S. 3471–3503.
- ^ Wolfgang Franke: Nutzplantenkunde. Nutzbare Gewächse der gemäßigten Breiten, Subtropen, und Tropen. 4. Auflage, Thieme, Stuttgart 1989. ISBN 3-13-530404-3
- ^ Lucia Impelluso: Die Natur und ihre Symbole. planten, Tiere und Fabelwesen. Parthas Verlag, Berlin 2005. ISBN 3-936324-03-4
- ^ Marianne Beuchert: Symbolik der planten. Insel Verlag, Frankfurt und Leipzig 2004, S. 279. ISBN 3-458-34694-5
Se også
[redigér | rediger kildetekst]Litteratur
[redigér | rediger kildetekst]- Wouter G. van Doorn og Uulke van Meeteren: Flower opening and closure: a review. Journal of Experimental Botany, Bd. 54, 2003, S. 1801–1812. Se teksten online (Afsnittet om blomsteråbning)
- Dieter Heß: Die Blüte. 2. opl., 1990, ISBN 3-8001-6434-5
- Special Issue: Major Themes in Flowering Research. Journal of Experimental Botany, Bd. 57, 2006 Se teksten online)
- Doug Soltis, Pamela Soltis og Jim Leebens-Mack: Developmental Genetics of the Flower. Advances in Botanical Research 44., 2006, ISBN 978-0-12-005944-7
- Guenther Theißen og Rainer Melzer: Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower. Annals of Botany, Bd. 100, 2007, S. 603–619. Se teksten online
Eksterne henvisninger
[redigér | rediger kildetekst]- Blomsterbiologi (pdf) Arkiveret 19. juli 2011 hos Wayback Machine (tysk)
- Blomster- og frugtbiologi Arkiveret 24. maj 2011 hos Wayback Machine (tysk)
- Blomsterbilleder fra Baseluniversitetets billedarkiv (tysk)
- Knop- og blomsterdannelsens fysiologi (tysk)
- Flower Anatomy Arkiveret 28. august 2005 hos Wayback Machine (engelsk)
- The Flowers Arkiveret 17. august 2005 hos Wayback Machine (engelsk)
| Søsterprojekter med yderligere information: |
| Botanikkens underemner |  | |
|---|---|---|
| Planter | ||
| Planteorganer | ||
| Planteceller | ||
| Planters livscyklus | ||
| Planters taxonomi | ||